#011: Characteristics of Fungi
What is a fungus? That question has been answered many ways over the years, reflecting an evolving understanding of this strange group of organisms. Initially, fungi were classified as a class in kingdom Plantae. As our understanding of the chemistry, life history, and evolution of fungi improved, Fungi was elevated to the rank of kingdom. Kingdom Fungi is placed in domain Eukarya and belongs to the major clade Opisthokonta. Opisthokonta also includes the kingdom Animalia, which makes animals close relatives of the fungi. But going beyond taxonomic definitions, what makes a fungus a fungus? It’s hard to answer that question because the fungal kingdom is very diverse and there are many similar organisms in other kingdoms. As a simple answer, Fungi are heterotrophic filamentous (or unicellular) eukaryotes with a chitin cell wall that reproduce by spores and digest their food externally.
Basic Features
If that last sentence didn’t make much sense, you’re not alone – even mycologists have a hard time figuring out what fungi are. Part of the problem is that fungi grow seemingly everywhere and therefore have adapted to many different environments, ecological roles, and life history strategies. The kingdom includes an estimated 2.2-3.8 million species (estimates range from 1.5 to 10 million species),1 so there are many opportunities for these organisms to adapt and evolve new and strange forms. Some fungi have become simpler over time, such as the unicellular Baker’s Yeast, while others have evolved to form complex multicellular structures like the mushroom Amanita muscaria. However, all these organisms have some basic features in common:
- Eukaryotic: Since fungi belong to domain Eukarya, they have a nucleus and membrane-bound organelles. Some bacteria form fungus-like chains of cells (ex: the actinomycetes), but they are not eukaryotes and do not have membrane-bound organelles. Fungi are one of the major branches of life that have evolved to be multicellular.
- Heterotrophic: Like animals, fungi cannot produce their own food and have to extract energy from organic matter in the environment. Fungi acquire food by decomposing dead organisms, by being parasites on living organisms, or by trading other resources for food in mutualisms such as mycorrhizas.
- Exoenzymes: Fungi digest their food externally by releasing enzymes into their surroundings. The enzymes break down complex molecules into simple ones such as sugars, which can then be easily absorbed by the fungus. Since these enzymes act externally, they are called “exoenzymes.” Fungi typically have a high surface area, which makes this process very efficient.
- Hyphae or Yeast: Fungi can either grow as hyphae or as yeast (see FFF#003), and some species switch between the two forms depending on the conditions. Hyphae are cylindrical chains of fungal cells that grow at the tip and can branch and fuse with each other to form networks. A network of hyphae is called a mycelium. Fungi that grow as hyphae are often called “filamentous.” The term “thallus” is used to refer to the complete mycelium. Yeast are unicellular fungi. Essentially, a yeast is a highly reduced hypha. Yeast sometimes form in chains, but they are still unicellular because no communication or coordination happens between the yeast cells.
- Non-motile: Fungi do not have structures like cilia, flagella, or pseudopods that drive movement in animals and other branches of life. Instead, they have to colonize new areas by growing into them or by being moved there by external forces. Some fungi, such as the chytrids (FFF#014) do have a short motile spore stage, but these spores are not the feeding part of the fungus. Chytrid spores have a single posterior whiplash flagellum that allows them to swim through the water. Animal sperm cells also have a single posterior whiplash flagellum – this shared feature was derived from a common ancestor and is a defining feature of the Opisthokonta (which literally translates to “rear flagellum”).
- Spores: Reproduction and dispersal in fungi is accomplished through spores. Spores are small (usually single-celled) structures that have the capacity to regrow an entire organism. Fungal spores can be haploid (the nucleus contains one set of chromosomes), diploid (the nucleus contains two sets of chromosomes), or dikaryotic (each cell contains two haploid nuclei) depending on where in its life cycle the fungus is. Spores are dispersed through environmental means, usually wind, water, or animal activity.
Cellular Characteristics
The basic features listed above are not enough to define Fungi. Some similar organisms like the oomycetes (FFF#054) share all of the basic features. However, they clearly differ at the cellular level. Any definition of fungi must therefore include some common cellular characteristics:
- Chitin: This is perhaps the most important cellular characteristic of fungi: they have cell walls that use chitin. Chitinous cell walls helped prove that oomycetes and slime molds (FFF#053) do not belong in the fungi. Fungi have cell walls that are primarily made of chitin (β 1-4 N-acetylglucosamine), but also include β-glucans and mannoproteins. Chitin is also found in the exoskeletons of insects and a very similar molecule – hyaluronic acid – is found in human skin (and many beauty creams). This molecule is another feature that links fungi and animals evolutionarily. Chitin is formed by linking molecules of acetylglucosamine into long chains. The chains tangle up with each other and become cross-linked, which gives the fungal cell wall its strength.
- Incomplete Nuclear Envelope Dissolution: In most Eukaryotes, in order for cell division to occur the membrane surrounding the nucleus has to dissolve. This does not happen in fungi. In fungi, the nuclear envelope partially dissolves, but does not completely disappear and the chromosomes never leave the confines of the nuclear envelope.
- Small Nuclei: Fungal nuclei are smaller than those of most other eukaryotes and generally can’t be seen under the light microscope. However, fungal cells usually contain several to dozens of nuclei, perhaps making up for the small size.
- Compact Genomes: One reason that fungal nuclei are small is that they don’t contain a lot of DNA. Fungi have small genomes – their chromosomes encode relative low numbers of genes – and lack long repetitive sections in their genetic code. Eukaryotic organisms typically have lots of space between their genes taken up by repetitive sections of DNA that don’t code for anything. These repetitive sections provide space for various DNA sequences that regulate gene activity. Fungi still have regulatory sequences, but the amount of space between and among genes and regulators is greatly reduced.
- Ergosterol: Cell membranes of fungi have ergosterol in them to keep them fluid. This compound is similar to cholesterol, which carries out the same function in animals.
- Lysine Synthesis: Fungi have a unique pathway to make the amino acid lysine. This is a useful feature when classifying an organism as a fungus; even the most bizarre fungi use this unique lysine synthesis pathway.
- Glycogen: Like animals, fungi store energy as glycogen, a long chain of sugar molecules. Plants store energy as starch, which indicates fungi are more closely related to animals than to plants.
Life Cycle
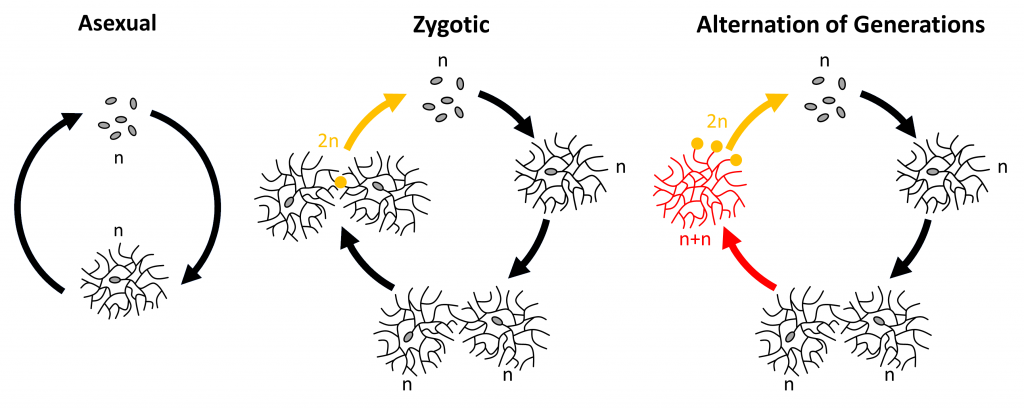
There are many variations of the fungal life cycle, but in general fungi use one of three life cycles: asexual, zygotic, or alternation of generations.
- Asexual: The asexual life cycle is the simplest. First, a spore lands in a suitable environment, germinates, and begins to grow. The fungus then produces spores through mitosis and the life cycle begins again. Asexual reproduction is found in all branches of fungi.
- Zygotic: The zygotic life cycle begins when a haploid spore lands in a suitable environment, germinates, and grows into a haploid mycelium. When a hypha from this mycelium encounters another sexually compatible haploid hypha, the two hyphae fuse. Two nuclei – one from each hypha – enter the point of fusion and then the nuclei fuse to create a diploid nucleus. This nucleus immediately goes through meiosis and produces sexual spores. The chytrids and zygomycetes (FFF#016) typically use a zygotic life cycle.
- Alternation of Generations: Once again, this life cycle begins when a haploid spore lands in a suitable environment, germinates, and grows into a haploid mycelium (the haploid generation). When a hypha from this mycelium encounters a hypha from a sexually compatible haploid mycelium, the two hyphae fuse. The two mycelia then become a single dikaryotic mycelium, with each cell containing two haploid nuclei – one from each parental mycelium (the dikaryotic generation). The dikaryotic mycelium continues growing. Eventually, some of the cells will convert to sexual cells. In the sexual cells, the nuclei fuse to become diploid and then undergo meiosis to produce haploid spores. Most ascomycetes (FFF#012) and basidiomycetes (FFF#013) use this life cycle.
Ecology
Fungi are some of the most important organisms on terrestrial earth, since they fill many ecological roles. Terrestrial ecosystems rely on fungi for cycling and extraction of nutrients, keeping populations in check by disease, and producing food in inhospitable environments.

- Decomposers: Many fungi, including the most familiar ones, are decomposers. Also known as “saprobes” or “saprophytes” (which can be modified to the adjectives “saprobic” and “saprophytic”), these organisms break down dead organic matter. Fungi play a major role in recycling nutrients in terrestrial ecosystems: decomposition frees up resources such as carbon, nitrogen, and phosphorous so they can be used by other organisms.
- Parasites: Parasites are organisms that feed on a living host. Fungi have evolved to parasitize many forms of life, from animals (including humans!) to plants to oomycetes, and even other fungi. Fungi excel at parasitizing plants and many significant plant pathogens are fungi.
- Mycorrhizas: Mycorrhizas are mutualistic symbioses between plant roots and fungi. The plants provide the fungi with energy in the form of sugar (produced through photosynthesis) and in exchange the fungi provide the plants with hard-to-get nutrients like nitrogen and phosphorous.
- Endophytes: This is a poorly defined ecological group, which is not surprising because it was established recently. Endophytic fungi live inside plant hosts without causing disease. Some endophytes support their host plants by producing chemicals that deter herbivores. Other endophytes are parasites that are kept at bay until the plant becomes stressed. Some endophytes seem to perform similar functions to mycorrhizas. But most endophytes serve an unknown function. So far, every plant studied has been found to harbor endophytes, making this an extremely important ecological group.
- Lichens: Lichens are “fungi that have discovered agriculture,” as lichenologist Trevor Goward puts it.2 A lichen is composed of a fungus and an alga. The alga uses photosynthesis to produce sugar for the fungus and the fungus provides structure and protection for the alga. The lichen relationship is often called a mutualism, but the fungus acts more like a parasite on the algae. This “controlled parasitism” is similar to the relationship between humans and agricultural crops. Lichens grow well in inhospitable places (such as bare rock) and produce energy using sunlight, which gives them unique roles in an ecosystem.
Divisions of Fungi and Taxonomy
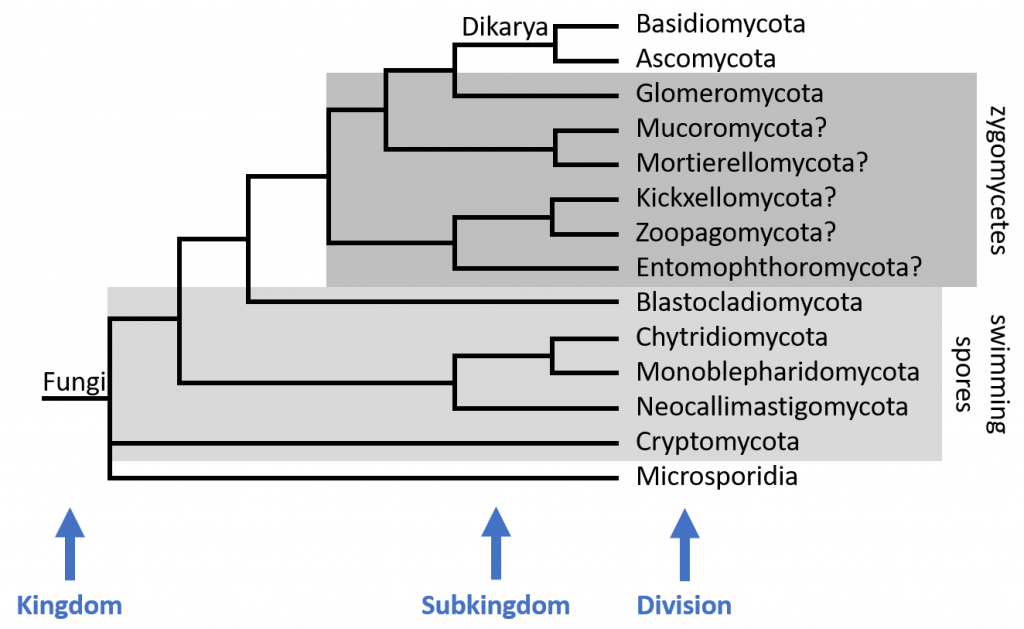
Fungal taxonomy has undergone many revisions in recent years, thanks to increasingly detailed genetic studies. Traditionally, Fungi was divided into four phyla (a taxonomic rank now called “division” in the Fungi) based on how they produce sexual spores: Ascomycota, Basidiomycota, Zygomycota, and Chytridiomycota.

Modern genetic studies have upheld the Ascomycota and Basidiomycota and demonstrated that the two divisions are closely related. Species in Ascomycota produce eight sexual spores inside a cell called an ascus, while the Basidiomycota produce four sexual spores externally on a cell called a basidium. Since Ascomycota and Basidiomycota are so closely related, mycologists place them in the subkingdom Dikarya. The name Dikarya reflects the fact that most species in these groups use an alternation of generations life cycle. Dikarya contains the most familiar fungi: mushrooms, Baker’s Yeast, and common molds, among others. Dikarya seems to be the most recently evolved lineage.3,4
The Zygomycota were split up multiple times and there is still disagreement over how they should be split up. It is clear that Zygomycota is not a real taxonomic group and it is no longer considered a division. Despite this, the term “zygomycete” (note the lowercase “z”) is still used by mycologists as a convenient morphological description. Zygomycetes form hamburger-like structures when they reproduce. When two sexually compatible zygomycete hyphae meet, they fuse together and each cell on either side of the point of fusion swells into a teardrop shape. Between these cells, a large circular spore called a zygospore forms. The nuclei from each hyphae fuse to become diploid and then divide mitotically to produce the multinucleate zygospore. The zygospore is thick-walled and is produced to survive adverse conditions. When the zygospore germinates, the diploid nuclei go through meiosis to return to a haploid state.5
The former zygomycete lineage with the most taxonomic support is the Glomeromycota (FFF#015). Most studies rank Glomeromycota as a division and place it as a sister group to the Dikarya. The Glomeromycota contain mycorrhizal fungi that reproduce only asexually. This is a very important group, since about 80% of land plants form mycorrhizas with the Glomeromycota.6
The rest of the zygomycetes are split up into two to eight divisions. There is not yet consensus over which lineages deserve to be ranked at the level of division, how the divisions are related taxonomically, or what the names of those divisions should be named. In the taxonomic tree below I have taken a middle ground, using five divisions: Mucoromycota, Mortierellomycota, Kickxellomycota, Zoopagomycota, and Entomophthoromycota. The “typical” zygomycete morphology is found in the Mucoromycota, which contains species like Mucor and Rhizopus that are commonly found in refrigerators growing on food. Entomophthoromycota contains insect pathogens, most notably the genus Entomopthora. Keep in mind, these groups are still not well understood, so the taxonomy of these divisions is likely to change.7–9
The Chytridiomycota in the old sense includes some of the earliest diverging fungi. These fungi have several methods for producing sexual spores, but none are very complex. Most of the species in this group produce spores with flagella, allowing them to swim in aquatic environments. The Chytridiomycota have also been split up, although Chytridiomycota has been retained as a division. Consensus seems to be emerging around four chytrid divisions: Chytridiomycota, Monoblepharidomycota, Neocallimastigomycota (FFF#018), and Blastocladiomycota (FFF#018).10,11 Chytridiomycota in the new sense includes species with swimming spores that are either saprobes or parasites. This division still contains the frog pathogen Batrachochytrium dendrobatidis (FFF#157). The division Monoblepharidomycota was split off of the core chytrids, but is still closely related to Chytridiomycota and shares many of the same features. Monoblephs produce swimming spores and are always saprobic.10 The division Blastocladiomycota was also split off from the old chytrids, so it also contains species with swimming spores. The Blastocladiomycota contains parasites, soil saprobes, and water molds, including the genus Allomyces, which is commonly used for research on chytrids.11 The division Neocallimastigomycota was established to contain obligate anaerobic fungi that live inside the guts of herbivores. These fungi are commensal – their hosts are neither harmed nor benefitted – and may have as many as 20 flagella to help move through the difficult environment of the animal gut.10
At the base of the fungal taxonomic tree are the divisions Cryptomycota and Microsporidia (FFF#017). These two divisions were recently added to the Fungi. The exact placement of Cryptomycota and Microsporidia has not been settled, but they both appear to have diverged from the rest of the fungi very early on in fungal evolution. Cryptomycota was established to contain Rozella species. Rozella lack cell walls and are intracellular parasites of fungi, oomycetes, and green algae. Several of the species produce spores with a single posterior flagellum.12 Microsporidia contains strange fungi with a highly reduced cell structure that live as intracellular parasites of animals, usually in the gut. Some taxonomists consider Microsporidia a sister group to Fungi instead of a member of the kingdom. Either way, the Microsporidia evolved near the base of the fungal taxonomic tree.13
Former Fungi
Early classification systems included slime molds and oomycetes as fungi, but both of those groups have since be moved to other kingdoms. Both slime molds and oomycetes have cell walls made of cellulose. Additionally, motile slime mold spores have multiple whiplash flagella in various arrangements and motile oomycete spores have one whiplash flagellum and one tinsel flagellum. Finally, slime molds spend most of their lives as amoebae rather than hyphae or yeast. These differences clearly demonstrate that slime molds and oomycetes do not belong in the Fungi or even in the Opisthokonta. Further research has revealed that both groups lack many of the basic features or cellular characteristics of fungi.14,15 The oomycetes are an interesting example of convergent evolution, since they evolved hyphae and tip growth (and the cellular structures associated with tip growth) independently of the fungi. So similar in appearance are oomycetes to fungi that oomycetes have actually been used as model organisms to study hyphal growth in fungi!16
The Origins of Mycology
Modern mycology begins after Carl Linnaeus (1707-1778, abbreviated to L. after scientific names) devised a system to scientifically name organisms. Linnaeus did not know fungi well, so although he named many mushrooms his classifications of fungi were not very useful.17 Systematic fungal taxonomy really began with Christian Hendrik Persoon (1761-1836, abbreviated Pers.), who developed a method of categorizing fungi based on spore-producing structures. Interestingly, Persoon seems to have never held a job, nor did he inherit substantial family money. Yet somehow, he still managed to eat, study fungi, and publish his work.18 The first professional mycologist was Elias Magnus Fries (1794-1878, abbreviated Fr.), who devoted his time to classifying fungi and is known as “the Linnaeus of Mycology.”19

It is important for American mycologists to relate the histories of mycology and America. Scientists (in Europe) did not begin naming fungi in earnest until well after the American Revolution (1776). Fries did not become a full faculty member until 1834.19 Around the same time, the American government systematically removed Native Americans from their ancestral lands in the eastern United States, a process that intensified after the Indian Removal Act of 1830. This act resulted in the deaths of thousands of Native Americans, the most well-known example being the Trail of Tears.20 American mycology began in this context, where Native Americans had been excluded from America.
One of the earliest American mycologists was Charles Horton Peck (1833-1917), who named about 3,000 American mushrooms between 1867 and 1913.21 Around the same time, Charles McIlvaine (1840-1909) began testing the edibility of hundreds of American mushrooms by eating them and became known as “Ole Ironguts.”22 These two men demonstrate that American mycology started from scratch – Native American knowledge of mushroom species and edibility has been absent from American mycology since the beginning. Our knowledge about mushroom edibility essentially begins in 1881, when McIlvaine first tried eating unknown mushrooms.22 Compare this to Europe and Asia, where traditions of eating mushrooms or using them medicinally extend back thousands of years (I know very little about mushroom traditions in Africa, Australia, and South America, so I cannot make comparisons with those regions). This in part explains why debates about edibility and proper names for species are still so common in American mycology: the science and traditions have existed for only about 140 years.
See Further:
Below are some links to other articles introducing the Kingdom Fungi.
https://herbarium.usu.edu/fun-with-fungi/what-are-fungi
https://www.apsnet.org/edcenter/disandpath/fungalasco/intro/Pages/IntroFungi.aspx
https://www.anbg.gov.au/fungi/what-is-fungus.html
Citations
- Hawksworth, D. L. & Lücking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. The Fungal Kingdom 79–95 (2017) doi:10.1128/microbiolspec.FUNK-0052-2016.
- Lichen Biology and the Environment: The Special Biology of Lichens. http://www.sharnoffphotos.com/lichen_info/biology.html.
- Swann, E. & Hibbett, D. S. Basidiomycota. Tree of Life web project http://tolweb.org/Basidiomycota/20520 (2007).
- Taylor, J. W., Spatafora, J. & Berbee, M. Ascomycota. Tree of Life web project http://tolweb.org/Ascomycota/20521 (2006).
- James, T. Y. & O’Donnell, K. Zygomycota. Tree of Life web project http://tolweb.org/Zygomycota/20518 (2007).
- Redecker, D. & Schüßler, A. 9 Glomeromycota. in Systematics and Evolution: Part A (eds. McLaughlin, D. J. & Spatafora, J. W.) 251–269 (Springer, 2014). doi:10.1007/978-3-642-55318-9_9.
- Benny, G. L., Humber, R. A. & Voigt, K. 8 Zygomycetous Fungi: Phylum Entomophthoromycota and Subphyla Kickxellomycotina, Mortierellomycotina, Mucoromycotina, and Zoopagomycotina. in Systematics and Evolution: Part A (eds. McLaughlin, D. J. & Spatafora, J. W.) 209–250 (Springer, 2014). doi:10.1007/978-3-642-55318-9_8.
- Zygomycota. Wikipedia (2020).
- Spatafora, J. W. et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 108, 1028–1046 (2016).
- Powell, M. J. & Letcher, P. M. 6 Chytridiomycota, Monoblepharidomycota, and Neocallimastigomycota. in Systematics and Evolution: Part A (eds. McLaughlin, D. J. & Spatafora, J. W.) 141–175 (Springer, 2014). doi:10.1007/978-3-642-55318-9_6.
- James, T. Y., Porter, T. M. & Martin, W. W. 7 Blastocladiomycota. in Systematics and Evolution: Part A (eds. McLaughlin, D. J. & Spatafora, J. W.) 177–207 (Springer, 2014). doi:10.1007/978-3-642-55318-9_7.
- Letcher, P. M. & Powell, M. J. A taxonomic summary and revision of Rozella (Cryptomycota). IMA Fungus 9, 383–399 (2018).
- Han, B. & Weiss, L. M. Microsporidia: Obligate Intracellular Pathogens within the Fungal Kingdom. Microbiol Spectr 5, (2017).
- Beakes, G. W., Honda, D. & Thines, M. 3 Systematics of the Straminipila: Labyrinthulomycota, Hyphochytriomycota, and Oomycota. in Systematics and Evolution: Part A (eds. McLaughlin, D. J. & Spatafora, J. W.) 39–97 (Springer, 2014). doi:10.1007/978-3-642-55318-9_3.
- Stephenson, S. L. 2 Excavata: Acrasiomycota; Amoebozoa: Dictyosteliomycota, Myxomycota. in Systematics and Evolution: Part A (eds. McLaughlin, D. J. & Spatafora, J. W.) 21–38 (Springer, 2014). doi:10.1007/978-3-642-55318-9_2.
- Deacon, J. W. Fungal Biology. (Blackwell Pub., 2006).
- Carl Linnaeus (1707 – 1778). Mushroom: the Journal of Wild Mushrooming http://www.mushroomthejournal.com/greatlakesdata/Authors/Linnaeus17.html.
- Christian Hendrik Persoon (1761 – 1836). Mushroom: the Journal of Wild Mushrooming http://www.mushroomthejournal.com/greatlakesdata/Authors/Persoon21.html.
- Elias Magnus Fries (1794 – 1878). Mushroom: the Journal of Wild Mushrooming http://www.mushroomthejournal.com/greatlakesdata/Authors/Fries10.html.
- Indian Treaties and the Removal Act of 1830. Office of the Historian, Foreign Service Institute https://history.state.gov/milestones/1830-1860/indian-treaties.
- Charles Horton Peck (1833 – 1917). Mushroom: the Journal of Wild Mushrooming http://www.mushroomthejournal.com/greatlakesdata/Authors/Peck22.html.
- Smith, P. Charles McIlvaine, Pioneer of American Mycophagy. Smithsonian Magazine https://www.smithsonianmag.com/arts-culture/charles-mcilvaine-pioneer-of-american-mycophagy-90379529/ (2012).
- Celio, G. J., Padamsee, M., Dentinger, B., Bauer, R. & McLaughlin, D. J. Assembling the Fungal Tree of Life: Constructing the Structural and Biochemical Database. Mycologia 98, 850–9 (2006).
- Blackwell, M., Vilgalys, R., James, T. Y. & Taylor, J. W. Fungi. Tree of Life web project http://tolweb.org/Fungi/2377 (2012).







![#011: Characteristics of Kingdom Fungi [Archived]](https://www.fungusfactfriday.com/wp-content/themes/hueman/assets/front/img/thumb-small-empty.png)