
#075: Ectomycorrhizae
Ectomycorrhizae are mutualistic relationships formed between trees and fungal species in both the Ascomycota and Basidiomycota. The main difference between ectomycorrhizae (abbreviated EM or ECM) and arbuscular mycorrhizae (discussed last week) is that in ectomycorrhizae the fungus never penetrates the host plant’s cells.
EM formation requires a concerted effort by both the plant and the fungus. Plant roots normally grow faster than EM fungi can colonize them, so the plant has to intentionally slow down its root growth. The end result of this is clusters of short, highly-branched roots. These “short roots” are found at intervals along quickly-growing “long roots,” which cannot be colonized by EM fungi. The mycorrhizal association is functional only in actively growing roots just behind the root tip. As a result, short roots are relatively short lived (most last less than a year) and new short roots must constantly be formed to maintain EM. Because EM are the result of coordinated growth between the fungus and the plant, short root morphology and branching will be different for each fungus/plant pairing. One thing all short roots have in common is that they lack root hairs. I suspect that this is because the EM fungus makes root hairs functionally redundant. Root hairs are designed to increase surface area and thus nutrient uptake, but EM fungi do the same thing much more efficiently.
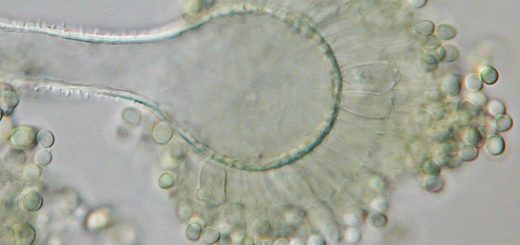
When an EM fungus finds a suitable host root, it encases the tip of that root in a thick layer of hyphae known as a “mantle.” Because of the mantle, short roots are noticeably thicker than other roots of the same order (number of branches from the main root), even at a low magnification. Short roots are completely surrounded by mantle hyphae, so all nutrients entering or exiting the root must go through the fungus. The mantle often persists after the short roots stop growing and the local EM association breaks down. In this case, the mantle is presumably acting as a storage structure.
The functional part of EM is found inside the root a few millimeters behind the root tip. Fungal hyphae growing between the root cells repeatedly branch and re-connect to form a net-like structure called a Hartig net, which envelops the cell. This net morphology serves to increase the surface area available for nutrient exchange. In angiosperms (flowering plants, including hardwood trees), Hartig net formation is limited to the epidermis, which is the outermost layer of root cells. Because these plants only make one layer of cells available to the fungus, the epidermal cells are often elongated and slanted to increase the EM surface area. In gymnosperms (conifers), the hyphae grow past the epidermis and form Hartig nets around cortical cells.
As with arbuscular mycorrhizae, EM fungi extend their hyphae into the soil. These soil hyphae (or “extraradicular hyphae”) serve to extract nutrients from the soil, transport nutrients, colonize new roots, and form reproductive structures.
EM fungi can be found in the phyla Ascomycota and Basidiomycota (and possibly in the family Endogonaceae from defunct phylum zygomycota). Several thousand EM-forming fungi have been identified within these groups, which dwarfs the total number of fungal species that form arbuscular mycorrhizae. Many of these fungi form mushrooms, and some are very well known. The Fly Agaric, truffles, morels (conditionally*), chanterelles, and boletes all form ectomycorrhizae. These fungi do not grow very well without their plant host(s), so no EM mushrooms have been successfully cultivated. Despite the wide diversity of EM fungi, the structures formed in EM associations are remarkably similar.
Only about 5% of plant species form EM. Despite this, EM is the most studied type of mycorrhizae, thanks to the presence of easily-observable fungal fruiting bodies. Plants tend to stick to one type of mycorrhiza, so most EM plants do not form arbuscular mycorrhizae. However, EM plants often form mycorrhizae with multiple species of EM fungi. In fact, the EM community beneath a tree can change over the course of that tree’s life. Certain fungi are known to prefer young hosts that are solitary or are at the forest’s edge while other fungi prefer old hosts in the middle of the forest. This phenomenon is known as ectomycorrhizal succession. The same principles can be found at work in other types of biological succession, but those usually describe species moving into a previously lifeless area. Most EM plants are trees found in temperate climates. The tropics, on the other hand, tend to be dominated by arbuscular mycorrhizal trees. Because of this, EM fungi appear to defy the rule that species diversity is centered in the tropics. Instead, the greatest diversity of EM fungi can be found in temperate forests. The plant families that contain a majority of EM species include: pine, oak, birch, willow, and walnut. For a more extensive list of EM plant species, see “Section D” on the mycorrhizas.info page linked below.
* Many morels appear to be saprobic and mycorrhizal at different points in their life.
See Further:
http://mycorrhizas.info/ecm.html
http://website.nbm-mnb.ca/mycologywebpages/NaturalHistoryOfFungi/Mycorrhizae.html






![#011: Characteristics of Kingdom Fungi [Archived]](https://www.fungusfactfriday.com/wp-content/themes/hueman/assets/front/img/thumb-small-empty.png)
4 Responses
[…] or in certain areas. This observation could explain the phenomenon of succession of EM fungi (see FFF#075 for more on EM […]
[…] mushroom hunters to tell boletes and agarics apart without picking them up. Most boletes are mycorrhizal with trees, so boletes are usually found growing on the ground around specific trees. Polypores […]
[…] One of the most commonly encountered mushroom morphologies is the polypore. Polypores are distinguished by a hymenium (spore-bearing surface) consisting of many, small pores. Unlike in boletes, the pore surface of the polypores cannot be easily separated from the rest of the mushroom. Polypores are a highly diverse group of mushrooms that come in a variety of shapes and sizes. The polypores you are most likely to encounter, especially during the winter, are tough to woody mushrooms that can last throughout the winter. Of these, the most common are the Turkey Tail (Trametes versicolor) and Trichaptum biforme. Both of these are medium-sized wood decomposers often found covering fallen logs or stumps. The tough fruiting bodies are annual, but can last for a year or more. Other polypores that last through the winter produce large, woody conks on the side of living trees or fallen logs. These mushrooms are perennial and get larger every year as new layers of pores are added. Some of the most common woody polypores include the Artist’s Conk (Ganoderma applanatum) and Fomes fomentarius. In the warmer seasons you can also find many fleshy polypores. These include the Chicken of the Woods (Laetiporus sulphureus), one of the easiest edible mushrooms to find and identify. Most polypores are wood decomposers, but a few fill other ecological roles. Fungi like Fomes fomentarius, Ganoderma applanatum, Grifola frondosa, and Inonotus dryadeus are parasitic to weakly parasitic on their host trees. Those in the genus Albatrellus are mycorrhizal. […]
[…] are two basic types of mycorrhizas: ectomycorrhizas (see FFF#075) and arbuscular mycorrhizas (see FFF#074). Most plants use arbuscular mycorrhizas and partner with […]