Developmental biology focuses on the question, “How do organisms start out as a single cell and then become large complicated structures with distinct tissues, shapes, and sizes?” In the context of mycology, studying development means researching the formation of structures such as mushrooms, sclerotia, and rhizomorphs. Not surprisingly, most developmental mycology research focuses on mushrooms, which are the largest and most interesting of the complex structures formed by fungi. In my previous post (FFF#234), I summarized what we know about mushroom development. Today, I will go into more detail on the mechanisms and processes that drive fruitbody development.
These topics are hard to discuss without going into the technical details, so I’ve provided a summary for each section with the technical details collapsed underneath. If you find any of these concepts particularly interesting, click on the button to expand the technical details. The two most important sections for understanding how mushrooms grow are “Cell Growth and Division” (under mechanisms) and the “Pattern Formation” (under processes).
Developmental Biology
Scientists studying developmental biology want to understand the sequence of events that structure large complex organisms. Researchers primarily study animal development, which can be applied to humans and used to understand human developmental problems (for example, birth defects and cancer). A wide range of animals are used in developmental biology, with fruit flies, nematodes, sea urchins, zebra fish, frogs, and mice being the most common. Surprisingly, these very different organisms use basically the same genes and strategies to control development.
Looking outside the animal kingdom, we see different genes are involved but the same strategies are used again and again. Because of this, developmental biology offers a useful framework for understanding development by looking at the strategies organisms use rather than specific genes or events. The major strategies found in all multicellular life are divided into five mechanisms and four processes. Mechanisms are the small-scale events that are used to carry out the processes: protein expression, cell-to-cell communication, cell growth and division, programmed cell death, and cell migration. Processes are overarching events that have to occur for development to be successful: pattern formation, morphogenesis, cell differentiation, and growth. By focusing on these mechanisms and processes, it is possible to compare and contrast widely different organisms.
Developmental Mechanisms
The five basic developmental mechanisms are tools used over and over again in different contexts to carry out all developmental tasks. Studying mechanisms takes a small-scale look at development: what is happening in a single tissue or a single cell or at a specific point in time to move development forward?
Protein Expression
Ultimately, what a cell does is determined by the proteins it produces. All the cells in your body contain the same genes, but they make different sets of proteins. For example, brain cells make proteins to conduct electricity, which are not needed elsewhere in your body. Because these electricity-conducting proteins are made and activated – “expressed” – specifically in your brain, your brain cells look different and have a special function compared to cells elsewhere in your body.
Flammulina velutipes has a slimy orange pileus and a tough black stipe. These differences are evidence of each tissue producing different proteins.
Fungi also change the proteins expressed in mushrooms in order to create different types of cells in different parts of the mushroom. This phenomenon is immediately apparent when you look at a mushroom. Consider Flammulina velutipes (FFF#071): the cap is orange and slimy, while the stipe is blackish and tough. All these physical differences are the result of differences in the proteins each tissue produces. The cap produces proteins that make orange pigments, while the stipe produces proteins for black pigments. The cap produces proteins that form slime. The stipe produces extra cell wall proteins to make the cells more rigid, resulting in a tough texture.
Studying how protein expression changes as the mushroom grows can tell us a lot about the cellular machinery used to build the mushroom. For more detail, expand the section below.
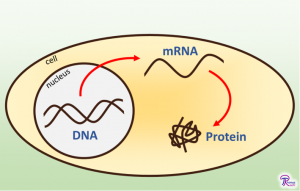
Proteins are built by reading instructions from genes, but there are three different parts to this process. First, the genetic instructions for each protein are stored in genes on DNA in the cell’s nucleus. DNA contains instructions for every protein a cell could make – called the genome – even if the cell isn’t making those proteins at the moment. Think of the genome as your computer’s hard drive: everything a computer can do is stored on the hard drive, but you never run all the programs at once.
Cells can’t directly read DNA, so an intermediate molecule is used to access the DNA information: mRNA. mRNA is built in the nucleus, where it copies down the instructions from genes in a process called “transcription.” It then exits the nucleus and goes into the cytoplasm – where those instructions can finally be turned into proteins. Think of mRNA as your computer’s RAM. RAM stores all the programs and files that a computer is currently using, such as the internet browser you’re using to read this post. To run any program, the computer copies the files off the hard drive and onto RAM, which allows the program to respond to your input. mRNA works in much the same way, since it copies down instructions in a format the cell can actually use. All of the mRNA molecules found in a cell is known as the “transcriptome” – essentially, it’s a list of all the programs the cell is currently running.
Proteins are responsible for all cell activity. To make a protein, DNA instructions are copied onto mRNA. Then, the instructions on mRNA are read and used to assemble a protein.
Finally, the cell has to read the mRNA instructions and create an actual protein. This is done at a structure called a ribosome using a process called “translation.” The ribosome reads the mRNA and assembles a chain of amino acid molecules as it goes along. Once the ribosome reaches the end of the instructions, the amino acid chain detaches and is then folded. A folded amino acid chain is known as a protein and this is the final structure that carries out a certain task within the cell. Proteins are like what you see on your computer’s screen. Your computer uses a program’s instructions (stored in RAM) to decide what color each pixel on the screen should be. The pixels are arranged in patterns that humans can interpret and interact with: the mouse cursor, text, buttons, etc. This is the whole purpose of the computer program: to provide you with information in a format you can use. Each pixel on the screen is similar to an amino acid, since it is carefully arranged to make useable patterns. The patterns are similar to the final proteins, since they are the part of the computer that you interact with. All of the proteins present in a cell – known as the “proteome” – determine what that cell actually does.
As a result of this three-part system to protein production, there are three ways to study protein expression: you can read the DNA, you can read the mRNA, or you can read the proteins. Reading the DNA tells you everything a cell can do, but it doesn’t tell you what the cell is doing at any given time. Because cells don’t use all their genes at once, protein expression is typically studied at later stages in the process. Looking at the transcriptome (mRNA) is a common method because reading mRNA is relatively easy. Some research also looks directly at the proteome (proteins), but that process is more difficult and expensive. Currently, transcriptome studies can provide more information and are therefore more common. The ability to read proteins is still progressing and will likely be a more common method in the future.
Hydrophobins are some of the most-studied proteins in mushrooms. These are short proteins divided into two halves: one half of the protein attracts water and the other half repels water. Hydrophobins are particularly important when hyphae are growing in the air. They coat the hyphae with the water-loving side against the cell to help keep water in and the water-repelling side facing out to form a barrier against the external environment.1,2 If you’ve ever tried to get a mushroom wet, you’ll notice that the water initially just bounces off the mushroom’s surface (especially in the area of the gills). This is because the hydrophobins coating the hyphae in the mushrooms repel water.
Because hydrophobins are important for aerial hyphae, these proteins are often specifically expressed in mushrooms. Agaricus bisporus (FFF#002), Schizophyllum commune (FFF#085), and Flammulina velutipes (FFF#071) all have specific hydrophobins that are produced only in mushrooms. These hydrophobins presumably help the mushroom maintain its structure – even gaps inside the mushrooms are coated with hydrophobins. Additionally, F. velutipes different hydrophobins for different tissues, including one (Fvh1) that is only produced by cells in the pileus.1–5
Cell-to-Cell Communication
Cells are constantly talking to their neighbors. This chatter serves many purposes and answers questions like, “Who are you?” “Who am I?” “Do you need food?” “Are we in danger?” and so on. Proper communication between cells is particularly important during development. For example, cell-to-cell communication ensures that all the cells in a particular tissue are making the correct proteins. It could also be used to keep all the cells in an organism growing at the same rate.
Cells in any organism have to communicate with each other in order to coordinate their actions.
Cell-to-cell communication in fungi has primarily been studied in the mycelium. We therefore know a lot about how fungal cells communicate during decomposition. Mycelium communication ensures that hyphae are evenly spread throughout the substrate to ensure the most efficient use of resources. In mushrooms, however, hyphae grow close together and therefore have to communicate differently.
Mushroom-specific cell-to-cell communication has not received as much attention. Most efforts have centered on identifying mushroom growth hormones. These studies used similar methods to studies on plant hormones, but unlike the plant research (which identified the hormones auxin, cytokinin, and gibberellin), mushroom hormone research did not produce any convincing results6. Expand the section below to read more about the evidence for mushroom hormones and related cell-to-cell communication topics.
Because of the structure of fungal cells, there are unique challenges for cell-to-cell communication in fungi. Fungal cells are arranged in hyphae: long cylindrical chains which are surrounded by a thick cell wall and grow at the tip. Even though hyphae are divided into cells using septa, the septa allow cytoplasm and most organelles to pass freely from one cell to the next. This makes it very easy to communicate between cells along a single hypha: the signal can quickly and easily travel through the septum to the next cell. This same hyphal structure makes it difficult to communicate between cells in adjacent hyphae: the signal must exit one cell, cross the cell wall, diffuse through the space between hyphae, cross the other cell wall, and then enter the other cell. Although hyphae do link with adjacent hyphae through anastomoses (see FFF#128), these connections are too infrequent to be responsible for all communication between hyphae.7,8 Consequently, fungal cell-to-cell communication exists in a very strange environment: sending a signal along one hypha can be done quickly, but sending a signal between two adjacent hyphae is very slow. This is different than both plant and animal cells, which are packed tightly together such that a signal going in any direction will take the same amount of time.
The only type of cell-to-cell communication studied extensively in mushrooms is hormonal communication. Hormones are compounds that act at a distance and cause specific cellular responses. In humans, compounds such as human growth hormone, estrogen, and testosterone are all examples of hormones: they are produced in specific places such as the pituitary gland (in the brain) but serve to coordinate growth throughout the body, particularly during adolescence. There is some good evidence that fungi also use hormones to coordinate development, but so far, no mushroom hormones have been definitively identified.
Stipe growth seems to be coordinated by hormones, at least in some species. In A. bisporus and F. velutipes, if you cut the cap off the stipe, the stipe will stop growing. Presumably, stipe growth ceases because the stipe grows in response to a signal from the cap. This signal is acting at a distance and therefore would seem to fit the definition of a hormone. However, attempts to find chemicals that impact stipe growth have produced inconclusive results.6,9,10
Other good evidence for hormones comes from A. bisporus and S. commune. If you extract liquid from A. bisporus mushrooms and apply the extract to S. commune mycelium, the mycelium will begin producing mushrooms. In this case, the A. bisporus mushrooms presumably contain some kind of signal hormone that promotes mushroom growth. S. commune apparently uses the same signal, which is why mushrooms start growing after the liquid is applied. Once again, attempts to figure out what the hormone was did not produce any useful information. As it stands, nobody has proven that mushrooms use hormones to communicate, even though mushroom growth seems to suggest the presence of hormones.6,11
Cell Growth and Division
A cell’s life cycle can be thought of in two phases: one phase where it gets bigger (cell growth) and another phase where it splits in half to make a copy of itself (cell division). In development, both cell growth and cell division can be specifically controlled to determine the size, shape, and number of cells in any given organ or tissue.
Determinate growth mushrooms are recognized by their ability to push aside debris and other mushrooms, as this Agaricus sp. is doing.
Fungi generally use either cell growth or cell division to enlarge their mushrooms. Some mushrooms – typically gilled mushrooms – get bigger by inflating all their cells (cell growth), which causes the mushroom to expand like a balloon (“determinate growth”). Other mushrooms – typically polypores – get bigger by adding new layers of cells to the outside of their fruitbodies (“indeterminate growth”). To produce the next layer, cells along the edge have to form new cells, which is done using cell division.
Almost all mushroom development research looks at mushrooms that use cell growth to get bigger. Expand the section below to read about these studies in greater detail.
In the typical model of mushroom development (used by determinate growth mushrooms), mushrooms get bigger through the mechanism of cell growth. This makes sense: if you make the cells inside a mushroom bigger, the mushroom itself will also get bigger. The process is similar to inflating a balloon. Mycologists have demonstrated this growth-by-cell-expansion using research termed “quantitative hyphal analysis” – a fancy way of saying “we measured how big the cells were.” Researchers measuring cells in Coprinopsis cinerea as it grew found that cells don’t always grow uniformly. In C. cinerea, the cells in the stipe start out tightly packed and all the same size. But in later stages, only the innermost ring of cells gets bigger. Because the cells in the very center and around the outside do not change size, they are drawn to the edges of the central ring. The end result is a stipe that is bigger but has a hole in the middle. By using cell growth in a specific area of the stipe, the mushroom both makes itself larger and changes its shape.6,12
The typical developmental model does not apply to all mushrooms, so the idea that cell growth drives mushroom growth does not work in every case. Unfortunately, cell growth/division has not been studied much outside of gilled mushrooms and we don’t really know what’s going on elsewhere.
Indeterminate growth mushrooms can be recognized by their ability to fuse with nearby mushrooms and grow around debris, as this Phellinus schweinitzii is doing.
However, we can get a pretty good idea just by watching mushrooms grow. If you watch polypores (for example, see Ganoderma lucidum in the movie Fantastic Fungi), you’ll notice that the old parts of the mushrooms stay put. Cut open an old polypore and you get the same impression: there are often rings inside that mark each day’s growth (or each year’s growth, for perennial polypores like Fomes fomentarius). These rings indicate that once a new layer forms, it doesn’t get bigger. In other words, the cells don’t grow after the layer is formed. Instead, the next layer must be formed by hyphae extending outward over top of the old layer. Because there are new hyphae forming, new growth must be driven by cell division. Indeterminate growth, therefore, is primarily driven by cell division.
Just by looking at cell growth and division, we come up with a simple mechanism that explains why determinate and indeterminate growth mushrooms behave differently as they grow. Determinate growth mushrooms primarily use cell growth, which means they inflate like a balloon and can push debris out of the way. On the other hand, indeterminate growth mushrooms use cell division to add new layers on top of old ones, which means they can grow around obstacles instead.
Of course, the indeterminate-growth-by-cell-division model is just a hypothesis, which should be tested by further research.
Programmed Cell Death
Organisms intentionally kill their own cells surprisingly frequently. This is a very orderly type of death where the dying cell divides itself into neat tiny packages, which are then consumed and recycled by nearby cells. One of the most prominent examples of cell death in humans is the webbing between our fingers. Human embryos initially grow fully webbed fingers (a relic of our evolutionary history), but before the baby is born the skin cells making up this webbing all kill themselves. The end result: unwebbed fingers that we can use to hold all kinds of tools. These types of events are called “programmed cell death” because the instructions for when and where the cells die are written into our DNA.
Inky caps use the mechanism of cell death to liquify their gills.
It’s actually quite difficult to study cells dying inside mushrooms, so I know of only two examples. Inky caps (FFF#177) use programmed cell death to liquefy their gills (called “deliquescence”): once spores are produced, the cells in the cap kill themselves, making the whole mushroom “inky.”13 Programmed cell death is also used in Agaricus bisporus (FFF#002) to create the gaps between the gills early on in mushroom development.14 You can read more about cell death in inky caps by expanding the section below.
One of the first people to study cell death was A. H. Reginald Buller (1874-1944)15, who is still widely known today for his careful observations of fungi. While watching the inky cap Coprinopsis cinerea grow, Buller noted that the gills liquefy beginning at the outermost edge and continuing toward the base of the gill.16 This is a specific pattern that happens every time the mushroom matures, demonstrating that the cell death is indeed programmed.
Later researchers delved more into the details of inky cap cell death and found that it is unusually messy. The cells of C. cinerea use enzymes to break down the cell walls around their dying hyphae, letting the cell contents spill out.13 Programmed cell death is usually designed to avoid creating a mess, but in the case of inky caps, creating a mess is the whole point. The inkiness – liquefied hyphal contents mixed with spores – is produced by specifically making the cells die in a very messy manner.
The purpose of liquefying the gills is to ensure efficient spore release. Spores mature from the outer edges inward, but it’s easiest to release spores on the outer edges because there’s nothing in the way. The easiest way to release spores on the inside would be if the outer edges just disappeared… which is exactly what happens in inky caps! By liquefying the outer edges, the inky caps can remove parts of the mushroom that have already released their spores. This ensures it is easy to release the spores near the center. Programmed cell death therefore helps shape the mushroom in a specific way to promote spore release.
Cell Migration
Sometimes, cells move. This can be very important for development, such as when a population of cells originates in one area but has to get to a different area to carry out its function. For example, during human brain development neurons literally crawl up a scaffold in the brain to form specific layers of tissues.17 The mechanism of cells moving (“cell migration”) therefore plays a vital role in proper brain formation. Of course, plant and fungal cells do not move – their rigid cell walls make that impossible. Consequently, cell migration is not a mechanism used in plant or fungal development.
Developmental Processes
The four major developmental processes are arguably more important than the various mechanisms. Processes use the mechanisms to define what the organism will look like, how big it will be, etc. These four big-picture processes occur in all complex multicellular life forms, making them useful for comparing development in distantly related organisms as well as closely related ones.
Pattern Formation
Proper pattern formation is vital to any complex organism. How did your body know where to put your head? Because early on, there was a pattern that determined which side of the embryo was up and which side was down. The process also involved patterns for left/right and front/back, as well as more complex patterns that were created on top of these: shoulder area, mouth area, eye area, etc. Cells pay attention to all these different patterns in order to figure out what kind of cell they’re supposed to be.
Pattern formation determines which parts of the mushroom will become each tissue. In this Amanita bisporigera, you can see the tissues already formed in the “button” on the bottom right.
In mushrooms, pattern formation occurs during the primordium. Amanita buttons are a good example of this phenomenon: cut one in half and you’ll find stipe, gill, and pileus tissues already present. By the end of the primordial stage, pattern formation is complete and the mushroom just has to expand upon those patterns.
The early steps in pattern formation have mostly been studied in gill tissue. Initially, gill tissue forms following a simple rule: “Where there is space, make gill.”*6 This results in a convoluted mess of tissue which is then straightened out by stretching the gills into shape.6 Expand the section below to read about the patterns responsible for creating gills.
By looking at tiny pieces of the early primordium under a scanning electron microscope, researchers discovered that early gills were convoluted wrinkles. A few steps are required to turn this mess into straight parallel gills. The first thing that happens is wrinkles form haphazardly. As more tissue forms, two or three new wrinkles can branch off from the initial ones. These first steps result in densely packed but randomly arranged gill tissue.18,19 Finally, the tissue has to be pulled into shape using mechanical stretching forces: either the pileus expands or the cells in the gills themselves inflate. In both cases, the wrinkles are subjected to stretching, which pulls them into neat rows.18,19 You can see the same phenomenon acting when you are folding a sheet and pull it tight from both sides. The sheet will stretch to form parallel lines extending from one hand to the other. Mushrooms use this mechanical process to arrange the wrinkles into regular linear ridges.6
After the basic pattern is established, that pattern is continued by the work of “organizing centers.”6 Organizing centers (another concept drawn from cell biology) are small areas around which patterns form – in other words, they are responsible for organizing groups of cells into patterns. The organizing centers for the gills are located between the gills near the gill base.6 The cells in that area direct cells on the right and left sides to become gill tissue. As this happens, the organizing center moves upward into the cap.20 Essentially, gill tissue isn’t added, but rather space between gills is excavated out of the pileus tissue. When organizing centers get too big (there is too much space between the gills), they split in half, which results in a new gill forming between two established gills.20 This process accounts for the existence of short-gills, but does not explain what causes gills to fork. Much more research needs to be done before we understand how these organizing centers form and are regulated.
A simple experiment demonstrates that gills do not extend from the tip: marking the gill margins with ink. In Volvariella bombycina and Pleurotus pulmonarius (see FFF#021), the marks remain at the bottom of the gill even as the gills get bigger.6,18 The tip, therefore, can’t be the area controlling gill formation. This agrees with previous research from C. cinerea that gills are organized from the base.6 These three species belong to three different families of gilled mushrooms, indicating that basal gill organization is widespread among agarics.
One odd feature of pattern formation in mushrooms is that it occurs multiple times throughout the fungus’ life cycle: in each mushroom and in each time mushrooms appear. This is unusual in the animal kingdom, where development typically happens once: the organism starts off as a juvenile and then becomes an adult and that’s it. A complete set of patterns formed once and only once in animals. Repeated pattern formation is common in plants, however: plants produce new leaves every spring and new fruits every summer, both of which require starting patterns anew. In this respect, fungal development is more similar to plant development (even though fungi are more closely related to animals).
Morphogenesis
The word “morphogenesis” literally means “creating shape.” This process focuses on how a ball of cells changes its shape to form various structures. In mushrooms, morphogenesis is mostly seen after the primordium stage. Consider an Amanita: all the tissues are packed into the egg-shaped primordium (“button”), but a fully grown Amanita mushroom is umbrella-shaped. At some point in its development, the mushroom’s shape had to change: this is the idea behind the process of morphogenesis.
A mushroom’s pileus margin can flip up when the gill cells grow faster than the cap cells.
Some good examples of morphogenesis in mushrooms come from measuring cells and from doodling on mushrooms. These experiments demonstrate that mushrooms primarily use the mechanism of cell growth to create their shape, but some shapes have to be created by cell division. For the details, expand the section below.
As we saw above, cell growth can make the whole mushroom larger. But, it is also used to drive morphogenesis. A detailed demonstration of this phenomenon comes from “quantitative hyphal analysis” (measuring cells) in Coprinopsis cinerea. The pileus of C. cinerea starts out egg-shaped. As the spores mature, the cap opens up to become conical and finally the edge flares up to make the cap bell-shaped. By measuring the cells in the cap, researchers demonstrated that this changing morphology is driven by cells in the cap growing at different rates. The cells at the very top of the pileus actually don’t get bigger at all, while cells in the gills expand greatly. Because the tissue underneath the cap gets bigger while the cap remains the same size, the gill tissue pushes the cap upward, eventually causing it to flare like a bell. In this way, expanding cells in particular places in the mushroom can be used to create a variety of shapes.
Cell growth clearly plays a large role in morphogenesis, but other factors can also be used to create shape. A good example of this is found in marking experiments on Pleurotus pulmonarius. When the gill edges of that species are inked, the marks stay near the center of the mushroom even as the cap gets bigger. The end result is gills where the inner (older) half of the gill is marked while the outer (newer) half of the gill is unmarked.6 This cannot be explained by simple cell expansion. If cell expansion were solely responsible for growth, the entire gill edge should have remained marked. Growth in P. pulmonarius, therefore, must be driven by cell division (adding new tissue) in addition to cell expansion. This use of growth by cell division is probably one reason why P. pulmonarius is fan-shaped instead of umbrella-shaped like most gilled mushrooms. Slight differences in the mechanisms used to drive morphogenesis are likely responsible for many of the diverse shapes we see in gilled mushrooms.
Cell Differentiation
In complex organisms, not all the cells are the same. You have skin cells, brain cells, muscle cells, etc., all of which look different and perform different functions. Cell differentiation is the process through which these differences were established. Cells within mushroom tissues also look different from each other and have specialized functions. These differences are primarily controlled by protein expression, which is the focus of a lot of current mushroom development research (see “Protein Expression,” above). However, microscopic work has also helped us understand how mushroom cells differ. Expanding the section below will provide you with more information on these microscopic studies.
Mycologists spend a lot of time staring through microscopes, and they apparently enjoy inventing fancy names for “looking at stuff.” One of those names is “hyphal analysis,” which really means “I looked at cells under a microscope and put them into categories.” Although this was most popular in the 1930’s, the categories are still useful today when identifying polypores and crust fungi. They are less useful from a developmental standpoint, but do provide one major insight: cells inside tissues can look different. Edred Corner classified hyphae into three different types: generative, skeletal, and binding hyphae.21,22 Each cell type is visually distinct and they are assumed to have distinct functions. At some point these cells became different from one another to perform specific roles in the mushroom. How those cells underwent differentiation has not been studied, but warrants further research by developmental mycologists.
One place where cell differentiation has been tracked in detail is in the gill tissue. For both Agaricus bisporus and Coprinopsis cinerea, mycologists have spent a lot of microscope time figuring out where each cell in the gill tissue comes from. In both species, the surface of the gill is composed of three types of cells: basidia (which produce the spores), cystidia (which are large and stick out of the gill surface for some unknown reason), and support cells (which provide structure). In A. bisporus, the support cells appear first and make up the entire gill surface early on. Later, some of the support cells get larger and differentiate into basidia and cystidia. All three types of cells come from the same original population of cells.23 In C. cinerea, cells once again from a layer over the gill surface. However, all of the cells in that layer develop into basidia and cystidia. Later on, cells from the inside of the gill grow into the layer of basidia and inflate to form the support cells. The three types of cells are therefore derived from two separate populations of cells.8 That the support cells form in such different ways is surprising since it indicates that different cell differentiation programs are used to form structures that carry out the same function. Does this mean that gills evolved separately in the two groups, or perhaps that forming support cells late is important for gill deliquescence? Answering questions like those has the potential to teach us about how gills evolved in both groups.
All mushrooms get bigger, although the process may involve slightly different mechanisms in different species.
Growth
The process of growth takes a large-scale look at how the organism becomes bigger. For mushrooms, growth is either a result of cell growth (the mushroom inflates like a balloon) or cell division (new layers of tissue form on the mushroom’s exterior). Both of those phenomena are discussed above under “Cell Growth and Division.” The process of growth was also discussed above under “Cell-to-Cell Communication” in the context of mushroom hormones. In addition to those two contexts, mushroom growth has been studied at the level of individual genes. Specifically, some genes have been identified that control stipe growth. For more details, expand the section below.
For people interested in the events controlling mushroom growth, the real question is what genes control growth and how they interact with each other. So far, several genes have been identified, but how they link together is still unknown. One of the most interesting genes is eln2, which encodes a cytochrome P450 in Coprinopsis cinerea. When eln2 is mutated, the stipe of C. cinerea won’t expand.24 Cytochrome P450s have a lot of roles in the cell, which probably means it’s just one step in a long pathway that controls stipe expansion. Having this as a starting point should help figure out the other genes that trigger and carry out stipe growth. Another mutation in C. cinerea to the gene ich1 prevents pileus formation. This gene appears to act in the nucleus, so it could be near the beginning or the end of the pileus formation pathway.25 One early gene involved in mushroom formation is pcc1, which can trigger mushroom production when mutated. The pcc1 gene interacts with mating type genes, which are a specific type of cell-to-cell signaling.26 These likely represent the first couple of links in the pathway that leads to mushroom formation, although how it all fits together is not yet understood.
One pathway that is well understood is the spore production pathway. These events are underpinned by meiosis, a special type of cell division used during sexual reproduction. Meiosis itself has been studied extensively, so it’s not surprising that spore production is well understood.6 It also helps that many mutant strains that fail to produce spores have been developed for C. cinerea27 as well as some cultivated mushrooms. By studying the point at which meiosis fails and the genes responsible for each failure, mycologists have been able to reconstruct the steps controlling spore formation.6 Because spore development focuses on individual cells instead of tissues, it doesn’t tell us a lot about mushroom development. Consequently, I won’t go into any more detail here, but there’s plenty of research out there if you want to learn more.
The Future of Developmental Mycology
It should be clear from reading the sections above that we know lots of pieces of the mushroom development puzzle, but very little about how those pieces fit together. Moving the field forward will require concerted efforts to research those links. Much of the recent research has focused on cell differentiation. Using new genetic tools, researchers are starting to figure out what makes tissues different from the level of individual genes. The more we learn about gene activity, the more we can start applying genetic tools to other areas such as pattern formation. Once a gene of interest is identified, it can be manipulated in coordination with microscopy to figure out when, where, and how that gene is active. Using systems like this, mycologists can finally start to work out the steps in mushroom development.
Models of mushroom development currently can’t explain why these boletes show features of both determinate and indeterminate growth.
In addition to cell differentiation, another good starting point would be pattern formation. Figuring out how patterns form would provide insight into the earliest steps in mushroom development. Additionally, it could help researchers finally move beyond gilled mushroom systems. Pattern formation must occur very differently in determinate and indeterminate growth mushrooms and describing those differences would help lay a framework for understanding multiple types of development. Such research would lead to a developmental model that could be applied beyond just gilled mushrooms.
This, in fact, is the biggest challenge for the field of mushroom development: applying models developed for gilled mushrooms to species with other morphologies. By reframing current knowledge in terms of developmental processes and mechanisms, mycologists can easily create a foundation for comparing development across species. As this post demonstrates, there is a wide array of research to draw from on each developmental topic, giving mycologists much to build on going forward.
Likewise, mycologists should also think about how developmental research applies to other areas of mycology, such as evolution, cell biology, and cultivation. There is so much we can learn about fungi by studying their fruitbodies and this area of research deserves more attention from mycologists.
A mycelium is a network of fungal cells that comprises the main body of the fungus.
Hyphae are cylindrical chains of fungal cells that can branch and fuse with each other to form networks.
The substrate is the material in which a fungus grows.
Cytoplasm is the liquid that fills cells. It is largely water, but also contains cell components and nutrients.
Organelles are structures inside cells, such as the nucleus (stores DNA), mitochondria (makes energy), and vesicles (store food, etc.).
Anastomoses are connections between nearby hyphae.
The primordium is an early stage in mushroom development during which tissue formation occurs. A complete primordium already contains the stalk, gills, and cap.
Agarics are mushrooms that release spores from gill-like structures under the cap (also called “gilled mushrooms”).
See Further:
All the information on human development was based on the developmental biology textbook I got for an undergraduate class on the subject (way back in 2012). If you want to learn more about animal development, reading a developmental biology textbook would be a good place to start.
For an even more in-depth look at mushroom development, see the book Fungal Morphogenesis by David Moore:
Wösten, H. A., Asgeirsdóttir, S. A., Krook, J. H., Drenth, J. H. & Wessels, J. G. The fungal hydrophobin Sc3p self-assembles at the surface of aerial hyphae as a protein membrane constituting the hydrophobic rodlet layer. Eur. J. Cell Biol.63, 122–129 (1994).
Lugones, L. G., Wösten, H. A. B. & Wessels, J. G. H. A hydrophobin (ABH3) specifically secreted by vegetatively growing hyphae of Agaricus bisporus (common white button mushroom). Microbiology (Reading, Engl.)144 ( Pt 8), 2345–2353 (1998).
Lugones, L. G., Bosscher, J. S., Scholtmeijer, K., de Vries, O. M. H. & Wessels, J. G. H. An abundant hydrophobin (ABH1) forms hydrophobic rodlet layers in Agaricus bisporus mushrooms. Microbiology (Reading, England)142, 1321–9 (1996).
De Groot, P. W. J., Schaap, P. J., Sonnenberg, A. S. M., Visser, J. & Van Griensven, L. J. L. D. The Agaricus bisporushypA gene encodes a hydrophobin and specifically accumulates in peel tissue of mushroom caps during fruit body development. Journal of Molecular Biology257, 1008–1018 (1996).
Ando, A., Harada, A., Miura, K. & Tamai, Y. A gene encoding a hydrophobin, fvh1, is specifically expressed after the induction of fruiting in the edible mushroom Flammulina velutipes. Curr. Genet.39, 190–197 (2001).
Moore, D. Fungal morphogenesis. (Cambridge University Press, 1998).
Williams, M. A. J., Beckett, A. & Read, N. D. Ultrastructural aspects of fruit body differentiation in Flammulina velutipes. in Developmental biology of higher fungi 333–351 (Cambridge University Press, 1985).
Rosin, I. V., Horner, J. & Moore, D. Differentiation and pattern formation in the fruit body cap of Coprinus cinereus. in Developmental biology of higher Fungi 333–351 (Cambridge University Press, 1985).
Gruen, H. E. Endogenous growth regulation in carpophores of Agaricus bisporus. Plant Physiology38, 652–666 (1963).
Gruen, H. E. Growth and rotation of Flammulina velutipes fruit bodies and the dependence of stipe elongation on the cap. Mycologia61, 149–166 (1969).
Rusmin, S. & Leonard, T. J. Biochemical induction of fruiting in Schizophyllum: isolation and preliminary purification of an inducing substance from Agaricus bisporus mushrooms. Plant Physiology61, 538–543 (1978).
Hammad, F., Ji, J., Watling, R. & Moore, D. Cell population dynamics in Coprinus cinereus: co-ordination of cell inflation throughout the maturing basidiome. Mycological Research97, 269–274 (1993).
Iten, W. & Matile, P. Role of chitinase and other lysosomal enzymes of Coprinus lagopus in the autolysis of fruiting bodies. Microbiology,61, 301–309 (1970).
Umar, M. H. & Van Griensven, L. J. L. D. Morphogenetic cell death in developing primordia of Agaricus bisporus. Mycologia89, 274–277 (1997).
Seaver, F. J. & Bisby, G. R. Arthur Henry Reginald Buller. Mycologia37, 275–277 (1945).
Buller, A. H. R. Researches on Fungi. vol. 3 (Longman Green & Co., 1924).
Budday, S., Steinmann, P. I. & Kuhl, E. Physical biology of human brain development. Front. Cell. Neurosci.0, (2015).
Chiu, S. W. & Moore, D. Development of the basidiome of Volvariella bombycina. Mycological Research94, 327–337 (1990).
Chiu, S. W. & Moore, D. A mechanism for gill pattern formation in Coprinus cinereus. Mycological Research94, 320–326 (1990).
Rosin, I. V. & Moore, D. Origin of the hymenophore and establishment of major tissue domains during fruit body development in Coprinus cinereus. Transactions of the British Mycological Society84, 609–619 (1985).
Corner, E. J. H. A Fomes with two systems of hyphae. Transactions of the British Mycological Society17, 51–81 (1932).
Corner, E. J. H. The Fruit-body of Polystictus xanthopus, Fr. Ann Botos-46, 71–111 (1932).
Allen, J. J., Moore, D. & Elliott, T. J. Persistent meiotic arrest in basidia of Agaricus bisporus. Mycological Research96, 125–127 (1992).
Muraguchi, H. & Kamada, T. A mutation in the eln2 gene encoding a cytochrome P450 of Coprinus cinereus affects mushroom morphogenesis. Fungal Genetics and Biology29, 49–59 (2000).
Muraguchi, H. & Kamada, T. The ich1 gene of the mushroom Coprinus cinereus is essential for pileus formation in fruiting. Development125, 3133–3141 (1998).
Murata, Y., Fujii, M., Zolan, M. E. & Kamada, T. Molecular analysis of pcc1, a gene that leads to A-regulated sexual morphogenesis in Coprinus cinereus. Genetics149, 1753–1761 (1998).
Kanda, T. et al. Isolation and characterization of recessive sporeless mutants in the basidiomycete Coprinus cinereus. Mol Gen Genet216, 526–529 (1989).
Thomas, this is such an excellent article. You have a great ability to explain complicated subjects such that a layperson can grasp what’s going on. Margaret (from MAW)
FFF#071: Flammulina velutipes is an edible mushroom that grows in two forms. In the wild, the umbrella-shaped mushrooms have a slimy orange pileus and a velvety stipe. When cultivated, the mushrooms become spaghetti-shaped with pale colors and tiny caps. Read More











![#140: Morchella angusticeps, the Black Morel of Eastern North America [Archived]](https://www.fungusfactfriday.com/wp-content/themes/hueman/assets/front/img/thumb-medium-empty.png)






![#011: Characteristics of Kingdom Fungi [Archived]](https://www.fungusfactfriday.com/wp-content/themes/hueman/assets/front/img/thumb-small-empty.png)
Thomas, this is such an excellent article. You have a great ability to explain complicated subjects such that a layperson can grasp what’s going on. Margaret (from MAW)
Thanks! I was a bit worried about this post because it does get rather complicated. I’m glad you were able to follow along!